
La révolution biotech des CRISPR

Quelques notions de biologie moléculaire
Les êtres vivants, observés au niveau cellulaire, se rangent en deux grandes catégories : ceux qui possèdent une cellule unique sans noyau, les procaryotes, qui comprennent les bactéries et les archées ; et ceux qui possèdent des cellules complexes à noyau, les eucaryotes, savoir les végétaux et les animaux, y compris l’animal humain. A noter que, bien que disposant d’éléments génétiques (molécules d’acides nucléiques, soit d’ARN, soit d’ADN), les virus ne sont pas, à proprement parler, des êtres vivants, puisqu’ils sont incapables de se répliquer par eux-mêmes. Par contre, ils se débrouillent très bien lorsqu’ils infestent une cellule-hôte. Ceux qui nous intéressent ici sont les bactériophages, qui colonisent les bactéries.
La cellule est l’unité fondamentale du vivant et, chez les eucaryotes, elle fonctionne de la même manière dans une plante ou chez l’être humain. Il existe donc une identité structurale des cellules animales et végétales. Le monde vivant est très diversifié, mais il obéit partout aux mêmes réactions biochimiques.
La cellule à noyau ou protoplasme se compose de deux grandes parties : le noyau, qui contient les chromosomes, autrement dit le patrimoine génétique de la cellule ; le cytoplasme, qui comprend le cytosol (milieu visqueux riche en protéines), dans lequel baignent les organites. On appelle organite tout élément cellulaire différencié assurant une fonction déterminée dans la vie de la cellule. Les organites qui nous intéressent ici sont les ribosomes, très nombreux, dont nous allons voir le rôle. Mais signalons au passage l’existence d’organites spécifiques aux bactéries, les plasmides. Ces plasmides possèdent un matériel génétique qui leur permet de se répliquer. En outre, ils peuvent être aisément construits par génie génétique pour des utilisations biotechnologiques (clonage du gène, séquençage de l’ADN…).
Le noyau, pour sa part, est constitué de 23 paires de chromosomes structurés en double hélice, comme l’ont découvert Crick et Watson en 1953. Les gènes sont portés par les chromosomes et occupent sur ceux-ci des emplacements fixes. Le séquençage du génome a précisément consisté à décrypter et à retranscrire cet ordonnancement des gènes sur les chromosomes.
Mais que fallait-il décrypter ? Les informations fines de chaque brin d’ADN. L’ADN (le code génétique) est constitué d’acides nucléiques, dont la brique de base est le nucléotide. Ces nucléotides, substances azotées basiques, n’existent que sous quatre formes dans la cellule : adénine (A), guanine (G), thymine (T) et cytosine (C). Le séquençage du génome consiste donc à lire la succession de ces quatre lettres, et c’est l’ordre dans lequel elles sont classées dans une séquence qui fixe la particularité du gène. On parle de séquence d’ADN car, sur un brin d’ADN, le code génétique est répété plusieurs fois, séparé par des zones dont on connaît encore mal l’utilité, et que l’on a baptisées un peu vite « ADN poubelle ». Il faut savoir que le code génétique ne représente que 2 % du matériel génétique total.
Les cellules se reproduisent en permanence, c’est la division cellulaire. Au cours d’une opération appelée « transcription », l’information contenue dans le noyau (l’ADN) va être copié sur une molécule, l’ARN messager, qui constitue une copie éphémère de l’ADN. ADN et ARN sont des chaînes d’acides nucléiques qui échangent de l’information chimique. L’ARN messager va être pris en charge par un ribosome, organite qui va jouer le rôle d’une tête de lecture. Les ribosomes, ou ARN ribosomique, sont également des acides nucléiques qui synthétisent les protéines en fonction des instructions reçues de l’ARN messager. Ces instructions sont fournies par une série de triplets de nucléotides adjacents (A, T, G ou C), triplets appelés codons. Chaque codon spécifie un acide aminé précis, sachant que le code génétique d’un chromosome est constitué de 64 codons qui peuvent produire 20 acides aminés différents. Les acides aminés jouent un rôle crucial dans le métabolisme, c’est-à-dire dans l’ensemble des transformations physico-chimiques qui interviennent dans les cellules. Ils sont au fondement de tous les processus vitaux tels que le transport et le stockage des substances nutritives. Ils agissent également sur les fonctions des différents organes.
Ces acides aminés se combinent entre eux pour former des peptides. C’est l’agencement de ces peptides qui va constituer les protéines, molécules fondamentales de l’organisme (20 % du corps humain). Parmi les protéines, très diverses, signalons le rôle des enzymes, qui catalysent les réactions chimiques.
Une protéine est construite à partir d’acides aminés qui sont assemblés par des ARN de transfert, lesquels s’assurent de la concordance des informations de la construction avec celles de l’ARN messager. Lors de la division cellulaire, chaque cellule réplique son ADN à l’identique.
Comment expliquer qu’un individu, qui est constitué de 50 000 à 100 000 milliards de cellules très différenciées, soit le produit de la division d’une seule cellule germinale, si les cellules se répliquent à l’identique ? Outre l’ARN messager, l’ARN ribosomique et l’ARN de transfert, il existe, au sein de la cellule, des ARN régulateurs, dont le rôle est de contrôler l’expression des gènes de structure, leur dictant quand et où s’activer. Ils orchestrent la vie de la cellule et conditionnent le bon fonctionnement de l’organisme. Ils jouent un rôle essentiel au stade de la différenciation cellulaire.
Ces rappels ne représentent qu’une approche grossière de la complexité du vivant. N’ont été évoquées que les notions nécessaires à la compréhension du sujet traité.
Le Paradigme du futur
Dès lors que l’homme a mieux connu le fonctionnement du vivant, il s’est empressé de vouloir le modifier. L’explosion des manipulations génétiques est en train de transformer radicalement de nombreux domaines, notamment la médecine, l’agriculture et l’élevage, et par conséquent l’avenir de l’espèce humaine, par exemple en produisant des animaux plus musclés pour obtenir plus de viande, des légumes plus gros et moins fragiles, des plantes plus résistantes et, surtout, des embryons humains génétiquement modifiés…
Depuis le 28 juin 2012, nous assistons à une révolution du génie génétique grâce à la publication de CRISPR/Cas9, un nouveau moyen, d’emploi facile et peu coûteux, de « modifier » l’ADN des cellules vivantes. Cette technique a ouvert la voie à d’autres, tout aussi révolutionnaires, telles que Cpf1, CasX et CasY. Dérivés d’un mécanisme de défense bactérien, ces outils de suppression et d’insertion de gènes se répandent dans les laboratoires de recherche. Il s’agit d’une technique qui vient s’ajouter à celles, jusqu’alors utilisées, pour couper l’ADN comme des ciseaux. Mais cette fois, l’outil est plus précis et plus rapide ; et il est plus aisé à manipuler. Depuis longtemps, et particulièrement depuis Gregor Mendel (1865), donc bien avant qu’il sache ce qu’est l’ADN, l’homme a croisé les espèces animales ou végétales pour sélectionner les expressions qualitatives qui l’intéressaient. La diversité des caractères repose sur l’apparition aléatoire et spontanée de mutations sur l’ADN. Les croisements appropriés ont pour but d’enrichir une population en individus porteurs de traits héréditaires d’intérêt (meilleure qualité, robustesse, productivité…). Qu’elles soient spontanées ou induites, les mutations par croisement de variétés sont aléatoires, nécessitant souvent des années d’expérimentation pour obtenir les caractéristiques souhaitées. Avec l’avènement du séquençage du génome de n’importe quel représentant du vivant, il devient possible d’en étudier les caractéristiques et, en utilisant des méthodes de manipulation génétique, d’y susciter les mutations désirées.
Jusqu’au début des années 90, les techniques utilisées pour obtenir des plantes ou des animaux génétiquement modifiés ne permettaient pas de s’assurer que la séquence d’ADN étranger serait introduite dans la zone ciblée, ce qui pouvait induire un effet délétère du transgène. La voie au développement des techniques de manipulation génétique a été ouverte par les travaux pionniers de Mario Capecchi, Martin Evans et Oliver Smithies, qui ont utilisé la recombinaison homologue dans des cellules souches embryonnaires pour insérer le fragment d’ADN de leur choix (transgène) à l’endroit désiré. Cependant, la fréquence de l’événement de recombinaison étant très faible, l’obtention d’organismes modifiés était longue et coûteuse. Des améliorations concernant l’efficacité de la stratégie et son extension à toutes sortes d’organismes ont été constamment apportées. Citons par exemple l’élaboration de « ciseaux moléculaires » très précis, les nucléases à doigts de zinc (ZFN, pour zinc-finger nuclease) ou encore les méga-nucléases, ainsi que les TALENs (Transcription Activator-Like Effector Nucléases), dont l’utilisation se répand dans les années 2010 ; ces techniques offrent la possibilité de couper la séquence d’ADN exactement à l’endroit choisi, et ce, dans le génome de nombreux organismes pluricellulaires. Ces coupures ciblées de l’ADN permettent ainsi de préparer le transfert des gènes d’intérêt – ceux qui présentent un caractère intéressant –, mais aussi d’augmenter considérablement les évènements de recombinaison, facilitant ainsi l’intégration du transgène. Les « ciseaux moléculaires » sont des enzymes de restriction, donc des protéines, qui ne peuvent couper qu’une seule zone dans une séquence génétique. Par conséquent, chaque gène visé impose la construction d’une protéine spécifique adaptée à la coupe souhaitée. D’où la nécessité de fabriquer les enzymes souhaitées et de tester leur efficacité. Pour faciliter la tâche des chercheurs, une société américaine, Addgene, met à leur disposition, sous forme de plasmides, une panoplie de 50 000 kits moléculaires différents, qui sont autant d’enzymes de restriction spécialisées selon les coupes à obtenir. On imagine la difficulté, pour chaque chercheur, d’identifier les bons outils dont il aura besoin pour mener à bien ses expériences. L’outil CRISPR va révolutionner tout cela.
Les séquences CRISPR ont été découvertes en 1987 lorsque le biologiste japonais Atsuo Nakata identifie, dans le chromosome de la bactérie Escherichia coli, des séquences d’ADN dont l’enchaînement des bases (A, C, T, G) se lit de la même manière dans les deux sens : à l’instar des mots « ressasser » ou « rotor », on parle de palindromes… Trois ans plus tard, les bio-informaticiens qui analysent et comparent ces séquences découvrent que les fragments d’ADN qui séparent ces palindromes ne sont pas d’origine bactérienne mais correspondent au matériel génétique de virus qui infectent les bactéries, les bactériophages. Ces séquences d’ADN, appelées CRISPR, sont présentes chez environ 50 % des bactéries et 90 % des archées connues. A noter que chaque bactérie ou archée dispose d’un CRISPR qui lui est propre, qui utilise une ou plusieurs protéines.
La fonction biologique naturelle de CRISPR ne sera finalement mise en lumière que vingt ans plus tard, en 2007, quand on constatera que les bactéries alimentaires résistent mieux aux bactériophages quand elles ont des séquences CRISPR que quand elles en sont dépourvues. Tout se passe comme si la bactérie gardait en mémoire la trace de la première infection virale en incorporant dans son génome un fragment du virus qui l’infecte, et qu’elle s’en serve ensuite pour se débarrasser de tout intrus viral déjà rencontré par elle ou par ses ancêtres. Rodolphe Barrangou, de l’Université d’État de la Caroline du Nord, et Philippe Horvath, scientifique principal chez DuPont, établiront et caractériseront le système de défense immunitaire bactérien CRISPR, à savoir l’existence d’une immunité adaptative des organismes unicellulaires. Dès 2012, les chercheuses Emmanuelle Charpentier, de l’Institut Max Planck de biologie des infections, et Jennifer Doudna, de l’Université de Berkeley, vont se servir du mécanisme précis par lequel le virus est éliminé, et impliquer une nucléase, l’enzyme Cas9 (CRISPR associated protein 9) exprimée dans de petits ARN, pour reconnaître la séquence virale visée et la couper à l’endroit souhaité. S’inspirant ainsi de cette réaction immunitaire bactérienne, elles vont avoir l’idée de la détourner pour en faire un véritable outil biotechnologique à la portée de tous, qui peut s’appliquer à n’importe quelle cellule, un outil qui permet de modifier à volonté le génome.
CRISPR/Cas9 a été employé sur le génome des mammifères dès 2013 par Feng Zhang, qui travaille pour le Massachusetts Institute of Technology. En avril 2015, des chercheurs du Centre Helmholtz et de l’Institut Max Planck, de l’Université d’Umeå, en Suède, ont découvert un nouvel auxiliaire de CRISPR, non plus l’enzyme Cas9, mais une autre molécule, le Cpf1, qui fournit un outil beaucoup plus précis et plus adapté au génome des mammifères. Les chercheurs ont passé ces dernières années à séquencer le génome des microbes qu’ils avaient recueillis dans des environnements variés. Le résultat de leur travail a été rassemblé dans une base de données compilant le génotype de ces micro-organismes au format terabase (1012 paires de base). En 2016, l’exploitation de ces données a permis de mettre en évidence le fait que les séquences qui contrôlent la protéine Cas9 sont également présentes dans le génome de deux archées, alors que, jusqu’à maintenant, celles-ci n’avaient été trouvées que dans les bactéries. Parce que les archées diffèrent biologiquement des bactéries, cette découverte d’une protéine Cas9 dans les archées offre un nouveau domaine d’étude intéressant pour la recherche CRISPR. Au même moment, cette même équipe a aussi découvert deux nouvelles protéines Cas parmi les bactéries, CasX et CasY.
Qu’est-ce que CRISPR
CRISPR est un guide ARN, une tête chercheuse que l’on peut coder à souhait pour cibler exactement le gène que l’on veut atteindre. En génétique, les CRISPR, ou Clustered Regularly Interspaced Short Palindromic Repeats (Courtes répétitions palindromiques groupées et régulièrement espacées), sont des groupes de séquences répétées à intervalles réguliers dans l’ADN des procaryotes (bactéries, archées). Il s’agit de séries de répétitions directes courtes et régulièrement espacées par des séquences appelées « spacer », généralement uniques, de 21 à 37 paires de bases.
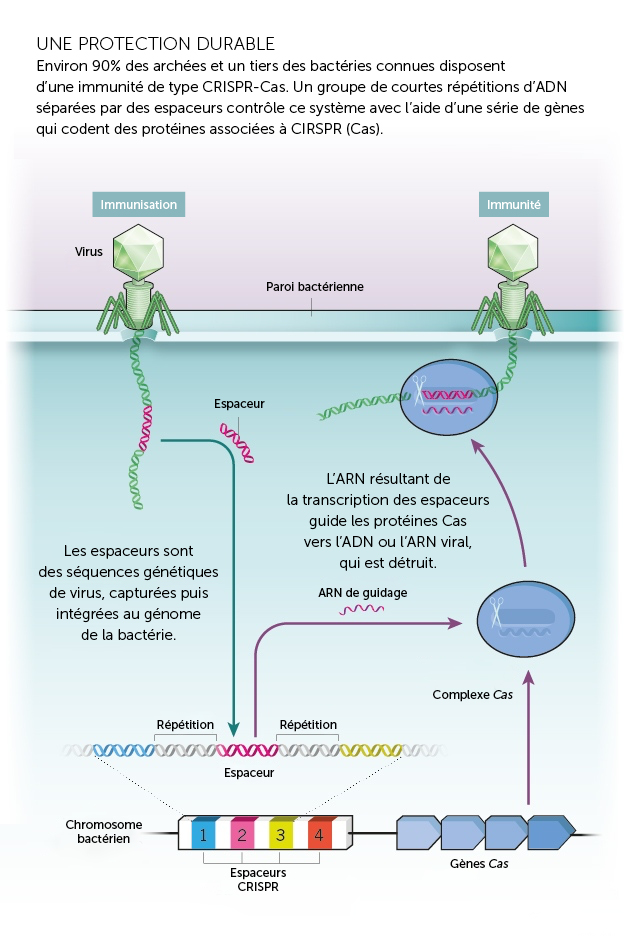 CRISPR est le mécanisme de défense naturel des procaryotes. Les procaryotes font face à l’assaut constant des envahisseurs génétiques que sont les virus. Ceux-ci sont dix fois plus nombreux que les procaryotes et tuent la moitié des bactéries présentes sur Terre tous les deux jours. Les procaryotes ont dû en conséquence développer un ensemble d’armes défensives pour faire face à la menace des virus. Chaque enzyme est programmée pour reconnaître certaines séquences, et un microbe est protégé seulement s’il a une copie du gène de l’agresseur. CRISPR s’adapte et se souvient des envahisseurs génétiques spécifiques d’une manière similaire à la façon dont les anticorps humains fournissent une immunité à long terme après une infection. C’est en étudiant les séquences entre les répétitions palindromiques qui semblaient correspondre à des génomes viraux, que les chercheurs en ont déduit la fonction de CRISPR. Certaines protéines associées au CRISPR ajoutent des séquences d’espacement au génome après que les bactéries et les archées ont été exposées à des virus ou plasmides spécifiques. Avec la ou les protéines associées, le système coupe et stocke l’ADN des envahisseurs. L’ARN fabriqué à partir de ces « espaceurs » dirige d’autres protéines pour absorber tout ADN ou ARN envahissant qui correspond à la séquence. Cela permet à l’organisme de conserver une mémoire génétique de cet envahisseur, qu’il peut alors référencer pour l’identifier rapidement s’il le retrouve à l’avenir.
CRISPR est le mécanisme de défense naturel des procaryotes. Les procaryotes font face à l’assaut constant des envahisseurs génétiques que sont les virus. Ceux-ci sont dix fois plus nombreux que les procaryotes et tuent la moitié des bactéries présentes sur Terre tous les deux jours. Les procaryotes ont dû en conséquence développer un ensemble d’armes défensives pour faire face à la menace des virus. Chaque enzyme est programmée pour reconnaître certaines séquences, et un microbe est protégé seulement s’il a une copie du gène de l’agresseur. CRISPR s’adapte et se souvient des envahisseurs génétiques spécifiques d’une manière similaire à la façon dont les anticorps humains fournissent une immunité à long terme après une infection. C’est en étudiant les séquences entre les répétitions palindromiques qui semblaient correspondre à des génomes viraux, que les chercheurs en ont déduit la fonction de CRISPR. Certaines protéines associées au CRISPR ajoutent des séquences d’espacement au génome après que les bactéries et les archées ont été exposées à des virus ou plasmides spécifiques. Avec la ou les protéines associées, le système coupe et stocke l’ADN des envahisseurs. L’ARN fabriqué à partir de ces « espaceurs » dirige d’autres protéines pour absorber tout ADN ou ARN envahissant qui correspond à la séquence. Cela permet à l’organisme de conserver une mémoire génétique de cet envahisseur, qu’il peut alors référencer pour l’identifier rapidement s’il le retrouve à l’avenir.
Qu’est-ce que Cas9 (CRISPR associated 9)
Dans la nature, le système CRISPR/Cas9 permet à la bactérie Streptococcus pyogenes de se défendre contre les virus. Le coup de génie, et sans doute la chance, de la chercheuse Emmanuelle Charpentier est d’avoir travaillé avec cette bactérie à protéine unique, l’enzyme Cas9, donc plus facile à mettre en œuvre.
Parmi les nombreux programmes CRISPR/Cas, le système de classe 2, qui utilise habituellement la nucléase Cas9, a été le plus étudié et, ces dernières années, il a été adapté pour être utilisé en biotechnologie. Le nom CRISPR/Cas9 s’applique aussi bien à la technique proprement dite qu’au matériel génétique introduit dans les cellules végétales ou animales lors d’une manipulation. En utilisant le système CRISPR/Cas9, il est possible de cibler des intervalles spécifiques de code génétique dans les organismes vivants, puis de couper une séquence pour modifier ces gènes plus précisément que jamais auparavant.
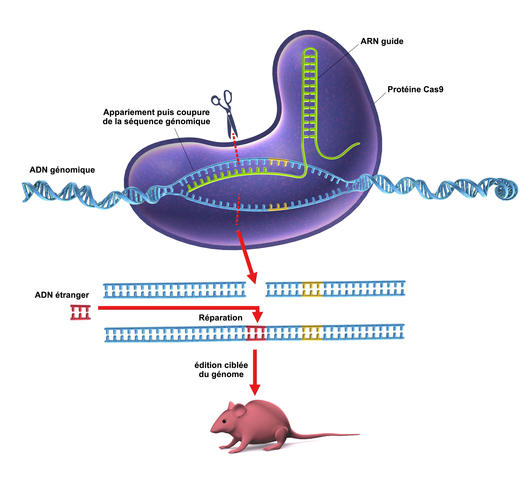
Le kit de modification contient un ARN « guide » (en vert sur le schéma) reconnaissant une séquence bien précise du génome (en bleu) ; qu’il transporte avec lui et ainsi guide, d’où son nom, le ciseau moléculaire Cas9 (la grosse protéine en mauve) à l’endroit ciblé. Une fois coupé, l’ADN peut être réparé ou modifié par insertion d’un fragment d’ADN approprié (en rose).
Cas9 est une endonucléase, c’est-à-dire une enzyme spécialisée pour couper l’ADN avec deux zones de coupe actives, une pour chaque brin de la double hélice. Celle-ci découpe l’ADN viral à une position déterminée par deux molécules d’ARN, nommées « CRISPR RNA » (crRNA) et « tracrRNA ». C’est un complexe formé de deux éléments : d’un côté, la molécule d’ARN (crRNA), de séquence homologue à celle de l’ADN que l’on veut exciser, et de l’autre, l’enzyme Cas9. Guidée par CRISPR, celle-ci effectue la reconnaissance de la séquence du génome viral par déroulement de son ADN et la vérification de complémentarité avec la région d’espacement longue d’une vingtaine de paires de base de l’ARN guide. Dans la cellule, la molécule d’ARN (crRNA) va reconnaître la séquence homologue sur l’ADN et s’y placer. L’enzyme Cas9 se charge alors de couper la chaîne ADN complémentaire à ce brin ARN. Le « trou » laissé par le passage du CRISPR/Cas9 pourra alors être comblé par n’importe quel nouveau fragment d’ADN. A cette construction génétique, vient s’ajouter un troisième élément, une molécule de « tracrRNA ». Cette séquence code un ARN qui s’hybridera avec l’ARN guide créé par la cellule par l’intermédiaire de CRISPR.
Parfois, l’ARN guide dirige l’enzyme vers une mauvaise partie du génome, ou vers d’autres répétitions des nucléotides qui n’étaient pas la cible visée par les chercheurs. Et bien que ces coupures hors cible ne se produisent pas souvent, elles pourraient avoir des effets désastreux sur l’organisme. C’est l’une des raisons pour lesquelles certains experts s’opposent à l’édition du génome humain.
Début 2016, les chercheurs ont modifié le nombre d’acides aminés que Cas9 utilise pour se lier à l’ADN. Après avoir testé une quinzaine d’itérations différentes, ils ont trouvé une version qui ne produisait pas d’effets détectables hors cible. Ils l’ont appelé SpCas9-HF1. Ajoutant plus d’acides aminés au mélange, les chercheurs ont découvert qu’ils pourraient étendre la portée de l’ADN que cible la protéine.
Les chercheurs ont officiellement reconnu 6 différents types de système CRISPR/CAS, avec 19 sous-types. « Et nous ne savons pas vraiment comment une fraction d’entre eux fonctionnent réellement » dit Luciano Marraffini, de l’Université Rockfeller à New York. Le déchiffrement de ces mécanismes pourrait constituer la clé de la recherche de nouvelles applications biotechnologiques pour les systèmes CRISPR/Cas.
Un « bouton STOP » pour CRISPR/Cas9
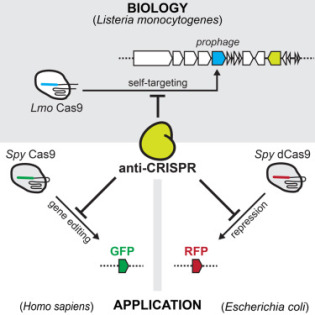 Si la technologie d’édition de gènes CRISPR est beaucoup plus facile à utiliser que les outils antérieurs, elle manque encore un peu de finesse. Parfois, elle modifie les cellules voisines que les scientifiques ne veulent pas changer, ce qui peut entraîner des résultats non désirés voir indésirables. Fin 2016, une équipe de chercheurs de l’Université de Californie de San Francisco a découvert que certaines protéines anti-CRISPR pouvaient protéger les cellules non ciblées contre les effets de l’outil d’édition génétique. L’équipe de chercheurs a examiné des souches de Listeria et évalué ses protéines pour trouver celle qui pourrait bloquer CRISPR. Pour Jospeh Bondy-Denomy, l’un des chercheurs, « ces inhibiteurs fournissent un mécanisme pour bloquer les applications CRISPR néfastes ou hors de contrôle, ce qui rend cette technologie plus sûre pour explorer toutes les façons dont elle peut être utilisée pour aider les gens ». Cela pourrait apaiser les critiques sur l’utilisation de cet outil et ouvrir la recherche sur la modification du génome humain.
Si la technologie d’édition de gènes CRISPR est beaucoup plus facile à utiliser que les outils antérieurs, elle manque encore un peu de finesse. Parfois, elle modifie les cellules voisines que les scientifiques ne veulent pas changer, ce qui peut entraîner des résultats non désirés voir indésirables. Fin 2016, une équipe de chercheurs de l’Université de Californie de San Francisco a découvert que certaines protéines anti-CRISPR pouvaient protéger les cellules non ciblées contre les effets de l’outil d’édition génétique. L’équipe de chercheurs a examiné des souches de Listeria et évalué ses protéines pour trouver celle qui pourrait bloquer CRISPR. Pour Jospeh Bondy-Denomy, l’un des chercheurs, « ces inhibiteurs fournissent un mécanisme pour bloquer les applications CRISPR néfastes ou hors de contrôle, ce qui rend cette technologie plus sûre pour explorer toutes les façons dont elle peut être utilisée pour aider les gens ». Cela pourrait apaiser les critiques sur l’utilisation de cet outil et ouvrir la recherche sur la modification du génome humain.
Qu’est-ce que Cpf1 (CRISPR from Prevotella and Francisella 1)
Certaines bactéries utilisent une autre enzyme, appelée Cpf1, afin de sectionner l’ADN étranger. Comme Cas9, Cpf1 est une endonucléase à ARN-guidée unique d’un système Cas de classe 2 ayant la propriété de couper les brins d’ADN de manière plus efficace. Cpf1 possède une caractéristique unique jusqu’à présent dans la famille d’enzymes Cas : elle coupe aussi bien l’ARN que l’ADN.
De plus, le système CRISPR/Cpf1 est une endonucléase plus petite et plus simple d’emploi que Cas9, permettant de dépasser certaines des limitations de CRISPR/Cas9 : elle ne nécessite aucune molécule supplémentaire pour transformer le pre-crRNA en CRISPR RNA (crRNA).
En outre, l’enzyme Cpf1 n’est guidée jusqu’à sa cible que par le crRNA, sans l’aide d’une molécule de « tracrRNA ». CRISPR/Cpf1 serait ainsi le système le plus minimaliste de cette famille décrit jusqu’à présent.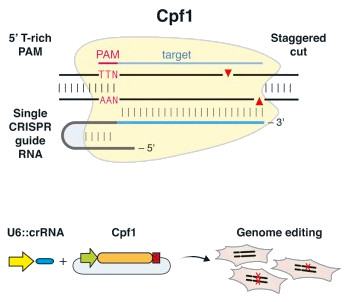
Le système CRISPR/Cpf1 consiste en une enzyme Cpf1 et un ARN guide qui clive l’ADN cible et positionne le fragment d’ADN à insérer au bon endroit sur la double hélice.
La mise en œuvre du système CRISPR/Cpf1 comporte trois étapes :
- L’adaptation : Les protéines Cas1 et Cas2 facilitent l’adaptation de petits fragments d’ADN dans le réseau CRISPR.
- Formation d’ARNc : Traitement d’ARN pré-CR produisant des ARNc matures pour guider la protéine Cas.
- Interférence : Le Cpf1 est lié à un crRNA pour former un complexe binaire pour identifier et cliver une séquence d’ADN cible.
Cas9 versus Cpf1
Associés à une protéine Cas9, deux molécules d’ARN sont nécessaires pour cibler et couper l’ADN quand Cpf1 n’en nécessite qu’une. Les deux protéines coupent l’ADN à différents endroits, offrant aux chercheurs de nombreuses options lors de la sélection d’un site d’édition. Cependant, Cas9 coupe les deux brins dans une molécule d’ADN à la même position, laissant les extrémités du brin ADN « émoussées » alors que Cpf1 laisse un fil plus long, créant des extrémités « collantes » qui sont plus faciles à utiliser. Cpf1 semble être plus en mesure d’insérer de nouvelles séquences sur le site de coupe, par rapport à Cas9. Bien que le système Cas9 puisse efficacement désactiver les gènes, il est difficile d’insérer des gènes ou de générer un knock-in (inactivation d’un gène pour le remplacer par un gène d’intérêt).
En résumé, les différences importantes entre les systèmes Cpf1 et Cas9 sont que Cpf1 :
- Dans sa forme naturelle, l’enzyme d’ADN Cas9 forme un complexe avec deux petits ARN, qui sont tous deux requis pour les activités de coupe. Le plus simple est CPF1 car il nécessite un seul ARN. Le CPF1 enzymatique est également inférieur à celui du SpCas9 standard, rendant plus facile son intégration dans les cellules et les tissus.
- De manière plus significative : Cpf1 coupe l’ADN différemment que Cas9. Lorsque le complexe Cas9 coupe l’ADN, il coupe les deux brins en laissant l’extrémité émoussée, entraînant souvent des mutations aléatoires lors de l’insertion du nouveau gène. Cpf1 coupe les deux brins en laissant des surplombs aux extrémités exposées. Cela devrait aider à l’insertion précise de morceaux d’ADN.
- Cpf1 coupe loin du site de reconnaissance, ce qui signifie que, même si le gène ciblé est muté sur le site de clivage, il est encore possible de le retailler, permettant de multiples possibilités pour un assemblage correct.
- Le système Cpf1 offre de la souplesse dans le choix de sites de destination. Comme Cas9, il doit d’abord se connecter à une courte séquence de l’ADN invasif ou PAM (Protospacer Adjacent Motif), et les objectifs choisis doivent être adjacents aux séquences naturelles du PAM. Le système Cpf1 reconnaît des séquences PAM très différentes de Cas9, donc propose de nouvelles zones d’intervention.
CasX et CasY (CRISPR associated X et Y)
Ces deux nouveaux systèmes CRISPR/Cas, CasX et CasY, utilisent des protéines plus petites que Cas9. Ces protéines sont composées respectivement d’environ 980 et 1 200 acides aminés. « Elles sont vraiment petites, surtout CasX ; cela signifie qu’elle est potentiellement plus utile », a déclaré Jillian Banfield. Celui-ci pense qu’il peut y avoir des composantes du système qui diffèrent, et que ces différences pourraient offrir de nouvelles informations pour améliorer les méthodes biotechnologiques actuelles. De même, Rotem Sorek, de l’Institut Weizmann des sciences en Israël, voit dans la plus petite taille des protéines CasX et CasY nouvellement découvertes une voie prometteuse, car « la livraison de petits gènes dans les cellules est beaucoup plus facile que la délivrance de gros gènes ». Cette recherche pourrait conduire à des innovations dans les techniques actuelles d’édition de gènes CRISPR.
Les « gene drives » (forçage génétique)
Les chercheurs s’interrogent sur les risques que pourrait faire courir le CRISPR en se déployant dans la nature au travers d’organismes modifiés. Une grande partie de l’attention s’est concentrée sur une méthode appelée « gene drive », ou « forçage génétique », qui peut répandre rapidement, à travers une population, un gène modifié. Les « gene drives » se nomment ainsi pour leur capacité à s’imposer à l’intérieur du génome des espèces modifiées génétiquement et à se répliquer.
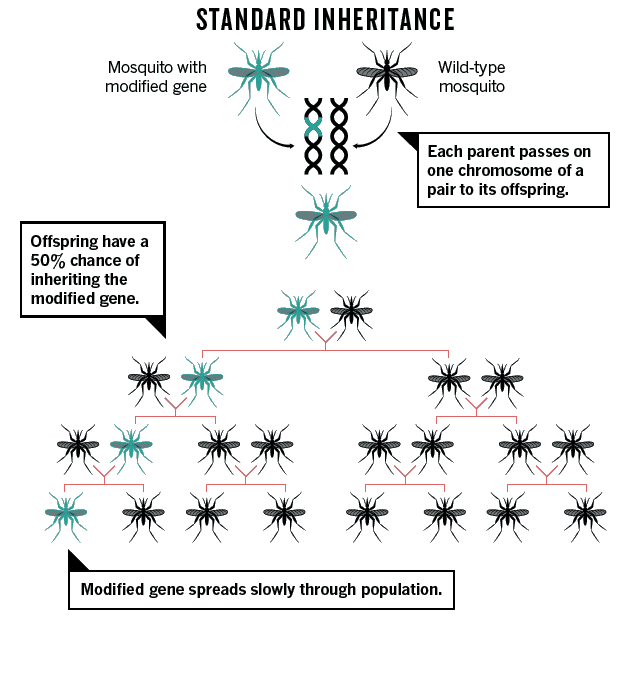
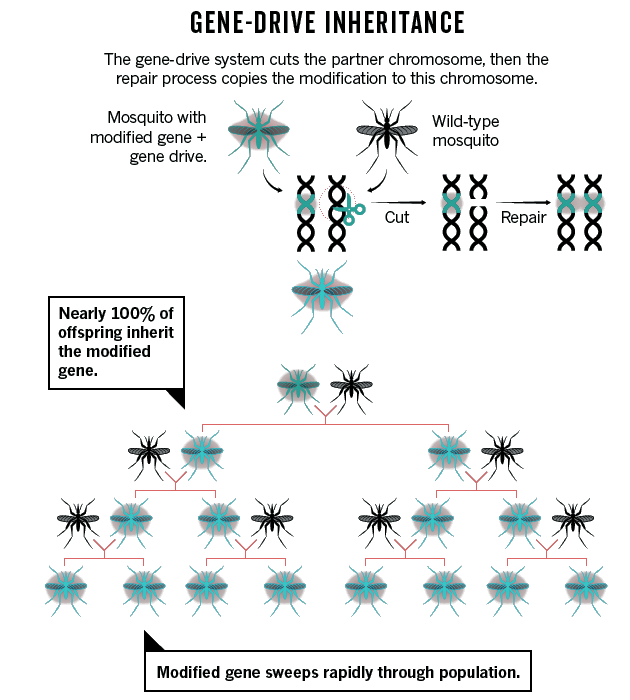
Ils amplifient les capacités d’expression des gènes auprès desquels ils se positionnent. Ils sont organisés naturellement de telle sorte qu’ils peuvent être transmis, lors de la reproduction de l’organisme, à la totalité des descendants de celui-ci, et non à la moitié d’entre eux. Cela signifie qu’ils vont se développer de manière plus rapide que la normale dans l’espèce traitée. En relâchant quelques individus qui possèdent une portion d’ADN modifié en laboratoire (appelée cassette « gene drive ») dans une population naturelle, on peut théoriquement obtenir, en quelques dizaines de générations, une population entièrement contaminée par la cassette « gene drive », dans laquelle une séquence spécifique d’ADN aura été introduite.
Ces modifications génétiques peuvent donc se répandre rapidement eu sein des populations, et peuvent devenir incontrôlables, car cette transmission se fait indépendamment des conséquences qu’elles peuvent avoir. Autrement dit, les « gene drives » sont transmis même s’ils ont des effets négatifs sur les espèces concernées. Des « gene drives » de synthèse ont été développés pour la première fois, dans le cadre des recherches en biologie génétique, afin d’être utilisés dans la lutte contre la reproduction des moustiques transmetteurs de la malaria. L’objectif était d’obtenir des populations de moustiques dépourvues des gènes transmetteurs de la malaria afin que celles-ci supplantent les populations originelles. Jusqu’ici, les espèces dites transgéniques, au génome modifié de manière traditionnelle, ne se répandent pas facilement dans la nature, car seulement 50 % de leurs descendants conservent les caractères introduits par manipulation génétique. Avec l’utilisation de « gene drives », ces caractéristiques sont maintenues dans 100 % des descendants. Or, utilisés au sein d’espèces mobiles, tels les moustiques ou les mouches du fruit, ces génomes modifiés pourront s’étendre à d’autres espèces, sans qu’il soit possible de prévoir ou prévenir leur dispersion. Donc cette pratique, si elle se développait, ne serait pas sans danger. Outre dans le domaine des moustiques, de nombreux chercheurs développent des « gene drives » synthétiques en vue de les diffuser au sein de diverses populations d’espèces sauvages afin d’altérer leurs capacités à transmettre des maladies, et plus simplement à empêcher la reproduction des espèces considérées – à tort ou à raison – comme nuisibles.
Quelles sont les applications des CRISPR ?
Ré-écrire l’Humain
CRISPR/Cas9 – appelé le « couteau suisse de la génétique » – fait miroiter d’incroyables débouchés pour la santé : soigner les cancers, mais aussi de nombreuses maladies génétiques comme la myopathie de Duchenne. Des expérimentations sont d’ailleurs en cours pour lutter contre la leucémie, la mucoviscidose, l’herpès, l’hépatite et même le VIH. D’autres s’orientent vers la recherche sur des traitements de la maladie d’Alzheimer. En neurosciences, plusieurs laboratoires utilisent déjà CRISPR/Cas9 pour dresser la carte d’identité génétique des maladies cérébrales (tumeurs, autisme…) ou pour étudier le développement des neurones. En procédant par inactivation successive des gènes, fragment par fragment, il devient possible d’identifier le rôle de chacun dans les maladies mentales, et donc d’établir le profil génétique de celles-ci. Pour l’autisme, par exemple, plus de 300 variations génétiques ont déjà été identifiées. Seulement, elles ne s’expriment pas toutes de la même façon au niveau des différents neurones. Avec CRISPR/Cas9, il devient possible d’étudier localement ces expressions génétiques. Selon une étude américaine, la tendance à boire en excès serait liée à un variant génétique. Des chercheurs ont identifié le rôle tout particulier joué par un gène spécifique, nommé KLB, qui « code » une certaine protéine qui lui est liée, nommée, elle, beta-Klotho. Cette protéine est l’un des constituants d’un récepteur situé dans le cerveau. Ce récepteur était déjà connu pour jouer un rôle dans la perception que nous avons de notre consommation de produits sucrés. Ce que l’étude met en évidence, c’est également son rôle dans la perception de la consommation d’alcool. En l’occurrence, le récepteur est censé réagir et inciter à la modération lorsque lui parvient, par l’intermédiaire du sang, une autre protéine (nommée FGF 21) qui est produite par le foie. Lorsque le gène KLB fonctionne bien, le cerveau perçoit le message envoyé par le foie et invite à la modération. Si le gène KLB est défaillant, il est possible de le remplacer en utilisant CRISPR, et donc de traiter l’addiction à l’alcool.
Re-modeler les animaux
Des chercheurs de Recombinetics, une société basée au Minnesota qui met au point des technologies génétiques pour l’agriculture et la biomédecine, ont inséré une version du gène POLLED dans le génome de cellules embryonnaires de bovins à un stade peu différencié. Ils ont ensuite utilisé le transfert nucléaire de cellules somatiques pour cloner les lignées de cellules génétiquement modifiées et implanter les embryons dans la vache/mère porteuse. Cinq veaux sont nés, dont deux sont encore en vie, Spotigy et Buri. Aucun des veaux n’avait de bourgeons de corne, ce qui suggère que la manipulation génétique a fonctionné. En introduisant ce gène qui élimine le « trait cornu », les chercheurs permettent d’éviter l’écornage des vaches laitières, une opération réalisée pour la sécurité des animaux et pour celle de leurs maîtres. Pour l’instant, les chercheurs n’ont noté aucun effet secondaire pour l’animal.
Usiner les plantes
Aux États-Unis, en avril 2016, un premier champignon modifié par CRISPR/Cas9 a été autorisé à la commercialisation. Ce champignon blanc commun (Agaricus bisporus) rendu résistant au brunissement est donc le premier végétal modifié génétiquement par CRISPR à recevoir son agrément de mise sur le marché. Le fait que ce champignon modifié génétiquement ne contienne aucun ADN étranger est à la base de cette décision. Actuellement, à l’échelle internationale, il n’est pas clairement établi si les plantes produites par les nouvelles techniques de génie génétique, dont CRISPR/Cas9, doivent être définies comme des OGM au sens des différentes réglementations en la matière, puisqu’elles ne sont pas différenciables des plantes cultivées de manière conventionnelle.
La recherche sur les modifications de gènes dans le monde végétal pourrait à terme améliorer la sécurité alimentaire, et permettre de fabriquer de nouveaux médicaments et de nouveaux vaccins à partir de plantes traitées par cette méthode, sous réserve de leur acceptation par la société. On se souvient que, en 2003, un champ de maïs traité pour soulager les enfants atteints de mucoviscidose a été saccagé dans le Puy-de-Dôme. Il en a été de même, dix ans après aux Philippines, s’agissant du riz doré, un riz enrichi en vitamine A pour vaincre les carences alimentaires dans le Tiers monde. Aujourd’hui, les opinions publiques sont effrayées par les potentialités des manipulations génétiques.
Quelles sont les enjeux éthiques de CRISPR ?
Aujourd’hui, manipuler les cellules germinales du génome humain serait prématuré, parce que ces techniques ne sont pas assez sûres. Elles peuvent introduire des modifications hors de la cible visée. Ainsi a-t-on pu constater que, chez l’animal, si on applique cette manipulation à un œuf juste fécondé, on peut produire des individus « chimères », car toutes les cellules ne vont pas porter les modifications souhaitées. On corrige quelque chose sans connaître tous les effets de cette correction, car les êtres vivants sont des systèmes hypercomplexes dont on connaît encore mal le fonctionnement. Mais les connaissances et la maîtrise de l’outil avancent très vite. En 2 ou 3 ans, les technologies CRISPR sont devenues de plus en plus sûres. Aussi, la puissance de l’outil commence à inquiéter. Avec la simplification des méthodes d’intervention sur le génome, de nombreuses tentations vont surgir. Les craintes se concentrent notamment sur la modification des embryons. Si l’on admet facilement la possibilité de réparer un gène défectueux chez un individu malade, est-on prêt à accepter de modifier le génome d’un enfant à naître pour lui incorporer des caractéristiques plus favorables (intelligence, force physique, beauté…) ? Où s’arrête la maladie et où commence le défaut purement esthétique ou social ? La Convention d’Oviedo de 1997 interdit l’utilisation d’embryons humains à des fins de recherche. Or, tous les pays ne l’ont pas ratifiée. Ainsi, en avril 2015, une équipe chinoise s’est-elle servie de CRISPR/Cas9 pour modifier le génome d’embryons humains. Les biologistes de l’équipe de Junjiu Huang (université de Sun-Yat-sen, Canton) ont travaillé sur des embryons humains non viables (car porteurs de chromosomes surnuméraires) et ont testé leur capacité à corriger une maladie redoutable du sang, la b-thalassémie. Même si une faible fraction seulement des 80 embryons traités avec le kit CRISPR/Cas9 a été corrigée, nul doute que, la technique s’améliorant constamment, le pourcentage de réussite ira croissant. Contrairement aux interventions effectuées après la naissance, qui ne modifient que ponctuellement l’individu, un changement génétique chez l’embryon concernerait nécessairement l’ensemble de ses cellules, y compris reproductrices (spermatozoïdes ou ovocytes) entraînant une transmission irréversible de la modification à sa descendance. C’est, comme le formulent déjà certains, la porte ouverte à l’eugénisme. Les Académies des Sciences de plusieurs pays (Angleterre, Chine, France, USA) se sont réunies à Washington début décembre 2015 pour débattre de l’utilisation biomédicale de CRISPR/Cas9. Elles ont reconnu la puissance et la facilité d’utilisation de cet outil pour traiter un grand nombre de pathologies d’origine génétique ou virale. Mais elles ont aussi noté que des améliorations techniques considérables étaient encore nécessaires avant d’envisager des essais cliniques. Ce qui pose le plus de questions n’est pas tant d’intervenir sur le génome des cellules somatiques (cellules musculaires, du sang, du foie…) que de corriger les cellules germinales et, partant, de modifier de manière irréversible le patrimoine génétique des générations à venir. Pour la première fois dans son histoire, l’humanité en en mesure de réécrire l’ADN de l’espèce, donc de modifier sa propre nature. Il faut prendre conscience du fait que l’on touche là à l’évolution de l’espèce humaine.
Terminologies
- CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) :
- regroupement de courtes paires de bases (21 à 48 – les valeurs changent selon les sources), répétition (jusqu’à 250 fois) de séquences d’ADN hautement conservées et (partiellement) palindromiques. Les répétitions ont en général une symétrie interne partielle, ce qui entraîne la formation de structures secondaires en épingle à cheveux.
- des séquences variables appelées espaceurs de longueur constante (20 à 58 paires de bases – les valeurs changent selon les sources) sont intercalées entre les séquences répétées.
- L’ensemble répétitions – espaceurs CRISPR est appelé matrice CRISPR (« CRISPR array » ou « CRISPR locus »).
- Cas (« CRISPR/associated ») : protéine codant les endonucléases associées à CRISPR
CRISPR/Cas est le premier (et jusqu’à présent le seul) système de l’immunité adaptative découvert chez les archées et les bactéries : celles-ci acquièrent ainsi une résistance à un ADN invasif.
Comme d’autres systèmes de défense, un système CRISPR/Cas est basé sur un mécanisme sophistiqué de discrimination [soi / non-soi].
Phase d’acquisition (aussi appelée d’adaptation) : un système CRISPR/Cas conserve de courtes séquences d’ADN invasif (les séquences espaceurs – « spacers ») comme éléments de mémoire du génome des virus et des plasmides rencontrés.
Phase de transcription – maturation : la matrice CRISPR (ensemble des séquences répétées – « repeats » – et des séquences espaceurs) est transcrite en un long transcrit primaire appelé pre-crRNA.
Celui-ci est maturé en un ensemble de courts fragments d’ARN appelés crRNAs (« CRISPR RNAs ») : chacun d’entre eux contient une séquence unique complémentaire d’un fragment d’ADN invasif.
Phase d’interférence : Les crRNAs sont utilisés comme ARN guide de la protéine Cas pour cibler et hydrolyser l’ADN apparenté de virus ou de plasmides invasifs lors de nouvelles infections.
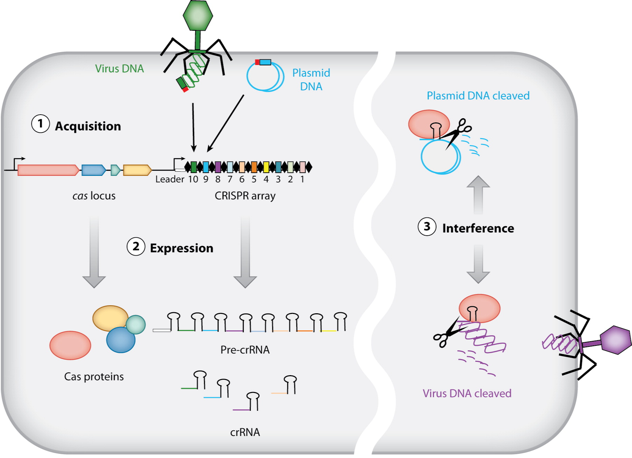
La séquence de tête (« leader sequence ») est une longue séquence riche en AT positionnée immédiatement en amont de la matrice CRISPR. Elle contient généralement à la fois le promoteur qui commande la transcription des crRNAs et la séquence de reconnaissance pour l’insertion de la séquence espaceur.
Matrice CRISPR :
- courtes séquences répétées (« crRNA ») qui constituent un élément capital de la régulation des systèmes CRISPR/Cas, car elles servent de point de fixation à la protéine Cas dans les trois phases du mécanisme.
- séquences espaceurs issues de l’ADN invasif
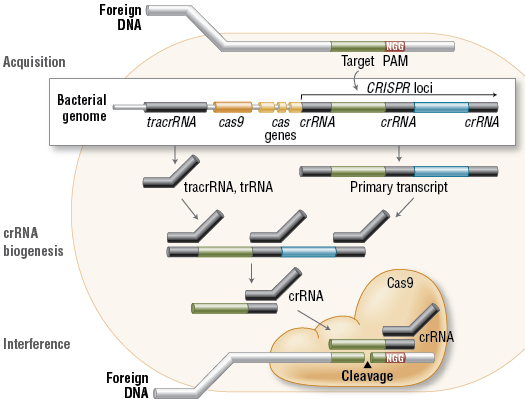 pre-crRNA : long transcrit primaire issu de la matrice CRISPR
pre-crRNA : long transcrit primaire issu de la matrice CRISPR
crRNAs : courtes séquences d’ARN issues de la maturation des séquences répétées du pre-crRNA
tracrRNA (« trans-acting small RNA ») : petit ARN qui s’apparie avec chaque répétition du pre-crRNA pour former un ARN double brin [tracrRNA:crRNA]
Séquence espaceur (porté par le crRNA mature) : séquence complémentaire de la séquence cible de l’ADN invasif (appelée proto-espaceur – « protospacer ») afin qu’il y ait hybridation.
PAM (« Protospacer Adjacent Motif ») : séquence de l’ADN invasif qui suit immédiatement la séquence cible proto-espaceur.
Bibliographie Sélective (MaJ 2025)
Alberts, Bruce, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts & Peter Walter (2015). Biologie moléculaire de la cellule (6e éd.). Bruxelles : De Boeck Supérieur.
Barrangou, Rodolphe (2017). CRISPR, la révolution génétique. Paris : Belin.
Barrangou, Rodolphe, Christophe Fremaux, Hervé Deveau, Marc Richards, Philippe Boyaval, Sylvain Moineau, Dennis A. Romero & Philippe Horvath (2007). CRISPR provides acquired resistance against viruses in prokaryotes. Science, 315(5819), 1709-1712.
Capecchi, Mario R. (2005). Gene targeting in mice: functional analysis of the mammalian genome for the twenty-first century. Nature Reviews Genetics, 6(6), 507-512.
Charpentier, Emmanuelle & Jennifer A. Doudna (2020). Le génie du gène : CRISPR, la révolution de l’édition génomique. Paris : Dunod.
Cong, Le, Feng Zhang, Patrick D. Hsu, Xuebing Wu, Wenyan Jiang, Luciano A. Marraffini & Jennifer A. Doudna (2013). Multiplex genome engineering using CRISPR/Cas systems. Science, 339(6121), 819-823.
Doudna, Jennifer A. & Samuel H. Sternberg (2018). Une révolution CRISPR : L’édition génomique et l’avenir de l’humanité. Paris : Quanto.
Evans, Martin J. & Matthew H. Kaufman (1981). Establishment in culture of pluripotential cells from mouse embryos. Nature, 292(5819), 154-156.
Girard, Michel (2018). CRISPR-Cas9 : Les ciseaux génétiques de la révolution génomique. Versailles : Éditions Quae.
Horvath, Philippe & Rodolphe Barrangou (2010). CRISPR/Cas, the immune system of bacteria and archaea. Science, 327(5962), 167-170.
Jacob, François (1970). La logique du vivant : Une histoire de l’hérédité. Paris : Gallimard. 2011
Jinek, Martin, Krzysztof Chylinski, Ines Fonfara, Michael Hauer, Jennifer A. Doudna & Emmanuelle Charpentier (2012). A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science, 337(6096), 816-821.
Lewin, Benjamin (2017). Les gènes (10e éd.). Bruxelles : De Boeck Supérieur.
Lodish, Harvey, Arnold Berk, Chris A. Kaiser, Monty Krieger, Anthony Bretscher, Hidde Ploegh, Angelika Amon & Matthew P. Scott (2017). Biologie moléculaire de la cellule. Paris : Dunod.
Mendel, Gregor (1996). Recherches sur les hybrides végétaux. Strasbourg : Presses universitaires de Strasbourg.
Smithies, Oliver, R. G. Gregg, S. S. Boggs, M. A. Koralewski & R. S. Kucherlapati (1985). Insertion of DNA sequences into the human chromosomal β-globin locus by homologous recombination. Nature, 317(6034), 230-234.
Watson, James D. & Francis H. C. Crick (1953). Molecular structure of nucleic acids: A structure for deoxyribose nucleic acid. Nature, 171(4356), 737-738.
Zetsche, Bernd, Jonathan S. Gootenberg, Omar O. Abudayyeh, Ian M. Slaymaker, Kira S. Makarova, Patrick Essletzbichler, Silvana E. Volz, Julia Joung, John van der Oost, Aviv Regev, Eugene V. Koonin & Feng Zhang (2015). Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell, 163(3), 759-771.


